With very few exceptions, mammalian antibodies of IgG type are composed of two identical heavy and two identical light chains linked to each other by disulfide bridges. Each chain consists of a species-specific constant region and a variable sequence that determines the antigen binding properties such as affinity and specificity (Amzel and Poljak, 1979) (figure 1).
Antibody preparations commonly used in scientific research are either polyclonal or monoclonal antibodies.
Polyclonal antibodies comprise a mixture of antibody molecules that differ in their variable sequences and therefore in their antigen binding properties (figure 2). Polyclonal antibodies are generally obtained from sera of immunized animals (Benner et al., 1981; Wootla et al., 2014).
After animal immunization, activated B-cells differentiate into plasma cells that finally produce huge amounts of soluble antibodies. Each of these highly specialized cells carries the genetic information for the secreted antibody (Benner et al., 1981; Wootla et al., 2014).
For the harvest of a maximum of antibody containing serum, the host animal is usually sacrificed and the plasma cells including the genetic information of the specific antibodies are lost.
Thus, the obtained antiserum represents a finite resource. New antiserum can only be obtained by re-immunizing more animals, which inevitably leads to certain batch variations no matter how thoroughly the new material is validated.
Monoclonal antibodies consist of identical antibody molecules that share the same antigen recognition characteristics (figure 3).
In 1975, hybridoma technology was introduced (Koehler and Milstein, 1975, Koehler and Milstein, 1976). In this method, antibody-producing B cells are fused with a myeloma cell line to generate so-called hybridoma cells, which can be propagated unlimitedly. Each hybridoma clone produces identical antibodies that share the same properties. The advantages of this technology are obvious. New antiserum batches no longer have to be extensively characterized and validated from scratch each time. Protocols no longer have to be laboriously adapted to changing antibody properties. Validation data that accumulate over time in the scientific community retain their validity and lead to an increasingly better characterized and valuable research reagent.
However, even monoclonal antibodies produced by hybridoma technology have certain limitations. They can undergo genetic drift which leads to undesired changes in specificity and affinity. In general, the genetic information of monoclonal antibodies is unknown and slight changes of antibody properties have to be minimized by careful cell-banking. Some hybridomas turn out to be unstable and stop expressing the antibody.
Figure 2: Polyclonal antibodies are a mixture of antibody molecules from the same species which are directed against the same target, but differ in their variable target recognition sequences.
Figure 3: Monoclonal antibodies are identical antibody molecules that share both the constant and the variable sequences.
Recombinant antibodies are monoclonal antibodies that are produced in vitro by using known antibody coding genes. The coding sequences of the heavy and light chain of an antibody can be obtained for example by hybridoma sequencing. The sequences are cloned into an expression vector and are transfected into a mammalian host cell line for antibody expression. Stably transfected cell clones can be generated and the expression can be adapted and optimized (Kuhnert and Reinhart, 2016).
Once the antibody is cloned, it is possible to modify it. The variable sequences can be mutated to alter binding specificity or affinity. The constant regions from one species can be replaced by constant sequences from another species, resulting in a chimeric recombinant antibody (Sahagan et al., 1986; Zalutsky et al., 1996) (figure 4). These genetically engineered chimeric recombinant antibodies retain all antigen recognition characteristics of the original antibody clone they originate from (figure 5).
They are produced under highly defined and controlled conditions as an infinite resource with minimal batch to batch variations. For the purification from cell culture supernatants no animals have to be sacrificed. The sequences are known and therefore the antibody can’t change due to genetic drift. Instead of that, there are multiple options for antibody engineering to adapt an antibody to specific experimental requirements. Synaptic Systems offers an increasing panel of chimeric recombinant antibodies with constant regions from rabbit, guinea pig, rat, human and chicken. They are all derived from our well performing mouse or rat monoclonal antibodies and can be combined with conventional species specific secondary antibodies. They are therefore valuable and reliable research tools with superior performance in your hands.
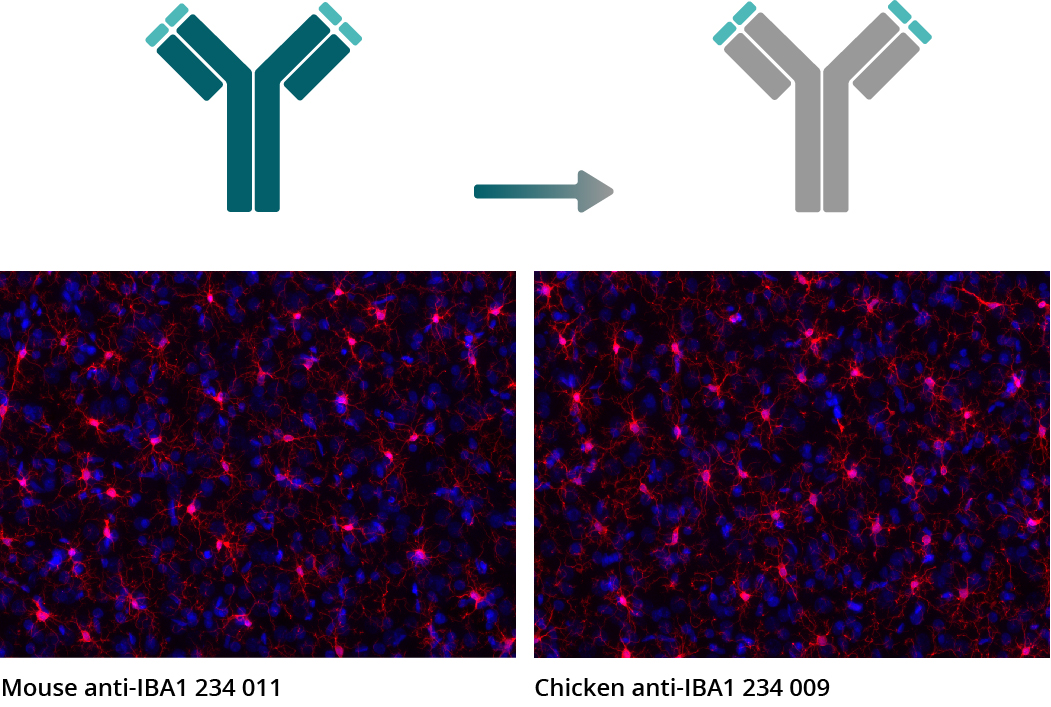
Figure 5: Chimeric recombinant antibodies retain all antigen binding properties of their original monoclonal antibodies as illustrated in representative pictures from comparative experiments. IHC images show immunostainings of rat brain sections obtained with the monoclonal mouse anti-IBA1 antibody 234 011 and the chimeric recombinant chicken anti-IBA1 antibody 234 009 derived from IBA1 antibody 234 011 (IBA1 = red, DAPI = blue).
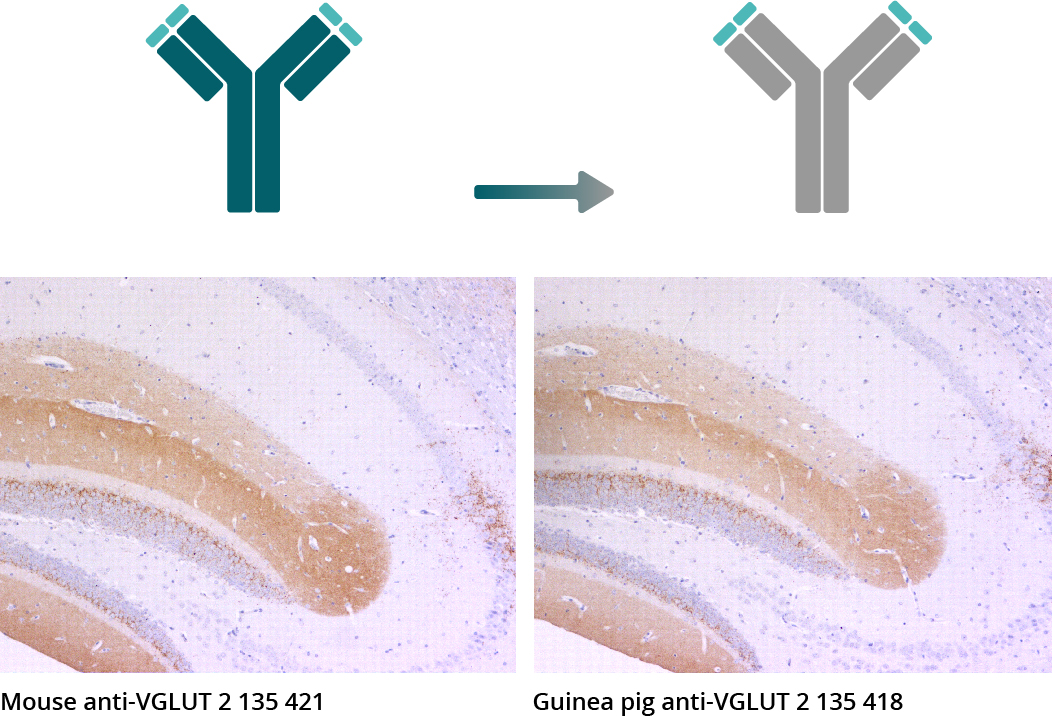
Figure 5a: IHC-P pictures illustrate indirect immunostainings of mouse brain sections utilizing the monoclonal mouse anti-VGLUT 2 antibody 135 421 and the emerging chimeric recombinant Guinea pig
anti-VGLUT 2 antibody 135 418 (VGLUT 2 = brown, haematoxylin = blue).
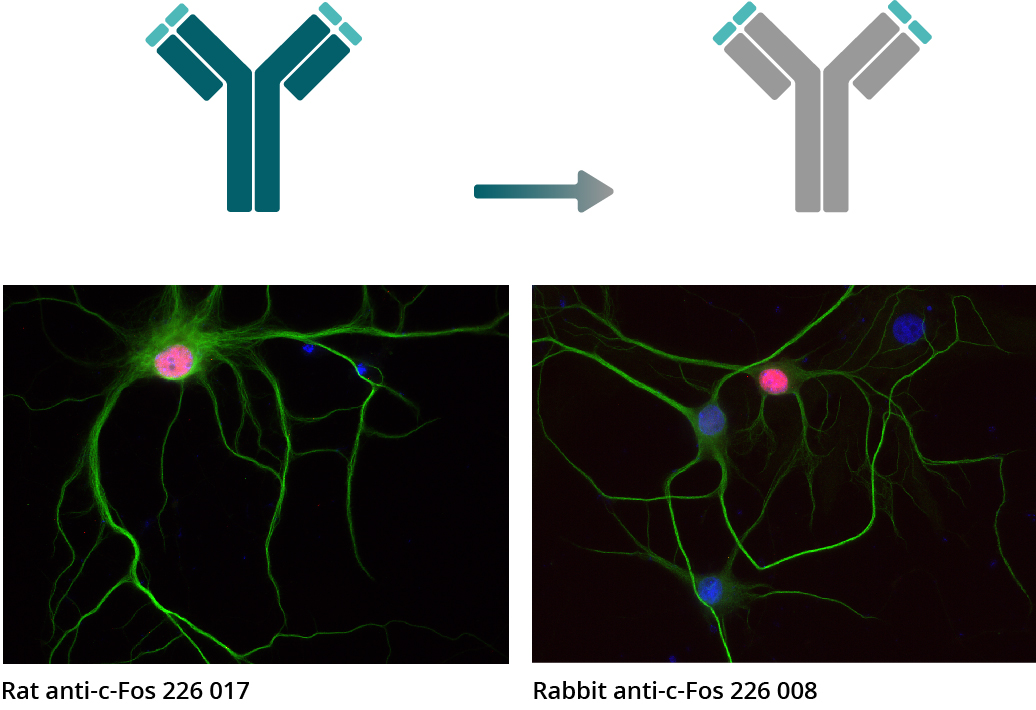
Figure 5b: In ICC experiments, c-Fos antibodies 226 017 (rat monoclonal) and c-Fos antibody 226 008 (rabbit recombinant) were used for indirect immunostainings of rat hippocampus neurons (c-Fos = red, MAP 2 188 002 (Polyclonal rabbit antiserum) and MAP 2 188 011 (Monoclonal mouse purified IgG) = green, DAPI = blue).
| Cat. No. | Product Description | Application | Quantity | Price | Cart |
|---|
| 310 118 | MLC-2V, rabbit, monoclonal, recombinant IgGrecombinant IgG | WB ICC IHC-P | 50 µg | $420.00 | |
| 310 308 | MLC-2V, Guinea pig, monoclonal, recombinant IgGrecombinant IgG | WB IHC-P | 50 µg | $420.00 | |
| 116 008 | Munc18-1, rabbit, monoclonal, recombinant IgGrecombinant IgG | WB ICC | 50 µg | $420.00 | |
| 260 008 | NET, rabbit, monoclonal, recombinant IgGrecombinant IgG | IHC | 50 µg | $420.00 | |
| 260 308 | NET, Guinea pig, monoclonal, recombinant IgGrecombinant IgG | IHC | 50 µg | $420.00 | |
| 266 008 | NeuN, rabbit, monoclonal, recombinant IgGrecombinant IgG | ICC IHC IHC-P | 50 µg | $420.00 | |
| 266 108 | NeuN, rabbit, monoclonal, recombinant IgGrecombinant IgG | ICC IHC IHC-P | 50 µg | $420.00 | |
| 266 308 | NeuN, Guinea pig, monoclonal, recombinant IgGrecombinant IgG | ICC IHC IHC-P | 50 µg | $420.00 | |
| 171 108 | Neurofilament H, rabbit, monoclonal, recombinant IgGrecombinant IgG | ICC IHC IHC-P | 50 µg | $420.00 | |
| 171 128 | Neurofilament H, rabbit, monoclonal, recombinant IgGrecombinant IgG | WB ICC IHC IHC-P | 50 µg | $420.00 | |
| 171 138 | Neurofilament H, Guinea pig, monoclonal, recombinant IgGrecombinant IgG | ICC IHC IHC-P | 50 µg | $420.00 | |
| 171 008 | Neurofilament L, rabbit, monoclonal, recombinant IgGrecombinant IgG | WB ICC IHC IHC-P | 50 µg | $420.00 | |
| 171 248 | Neurofilament M, rabbit, monoclonal, recombinant IgGrecombinant IgG | WB ICC IHC IHC-P | 50 µg | $420.00 | |
| 129 508 | Neuroligin2, rabbit, monoclonal, recombinant IgGrecombinant IgG K.O. | WB ICC IHC IHC-P | 50 µg | $420.00 | |
| 129 518 | Neuroligin2, Guinea pig, monoclonal, recombinant IgGrecombinant IgG K.O. | WB ICC IHC IHC-P | 50 µg | $420.00 |
Amzel and Poljak, 1979: Three-dimensional structure of immunoglobulins. PMID: 89832
Benner et al. 1981: Immunoglobulin isotype expression. II. Frequency analysis in mitogen-reactive B cells. PMID: 6975718
Wootla et al., 2014: Polyclonal and monoclonal antibodies in clinic. PMID: 24037837
Koehler and Milstein, 1975: Continuous cultures of fused cells secreting antibody of predefined specificity. PMID: 1172191
Koehler and Milstein, 1976: Derivation of specific antibody-producing tissue culture and tumor lines by cell fusion. PMID: 825377
Kuhnert and Reinhart, 2016: Advances in recombinant antibody manufacturing. PMID: 26936774
Shahagan et al., 1986: A genetically engineered murine/human chimeric antibody retains specificity for human tumor-associated antigen. PMID: 3088107
Zalutsky et al., 1996: Chimeric anti-tenascin antibody 81C6: increased tumor localization compared with its murine parent. PMID: 8832699