Cat. No.: 104 318
Amount: 50 µg
Price:
$420.00
|
|
|
|
| Cat. No. 104 318 |
50 µg purified recombinant IgG, lyophilized. Albumin and azide were added for stabilization. For reconstitution add 50 µl H2O to get a 1mg/ml solution in PBS. Then aliquot and store at -20°C to -80°C until use. Antibodies should be stored at +4°C when still lyophilized. Do not freeze! |
| Applications | |
| Clone | Gp69.1 |
| Subtype | IgG2 (κ light chain) |
| Immunogen | Synthetic peptide corresponding to residues near the amino terminus of rat Synaptobrevin2 (UniProt Id: P63045) |
| Reactivity |
Reacts with: mouse (P63044), rat (P63045), human (P63027), hamster. No signal: chicken, zebrafish. Other species not tested yet. |
| Specificity | K.O. validated |
| Matching control protein/peptide | 104-2P |
| Remarks |
This antibody is a chimeric antibody based on the well known monoclonal mouse antibody 69.1. The constant regions of the heavy and light chains have been replaced with Guinea pig specific sequences. The antibody can therefore be used with standard anti-Guinea pig secondary reagents. Important: Although the epitope is still present in the Botox B cleavage product (aa 1 - 76), it is not recognized, probably due to conformational masking of the epitope. |
| Data sheet | 104_318.pdf |
|
|

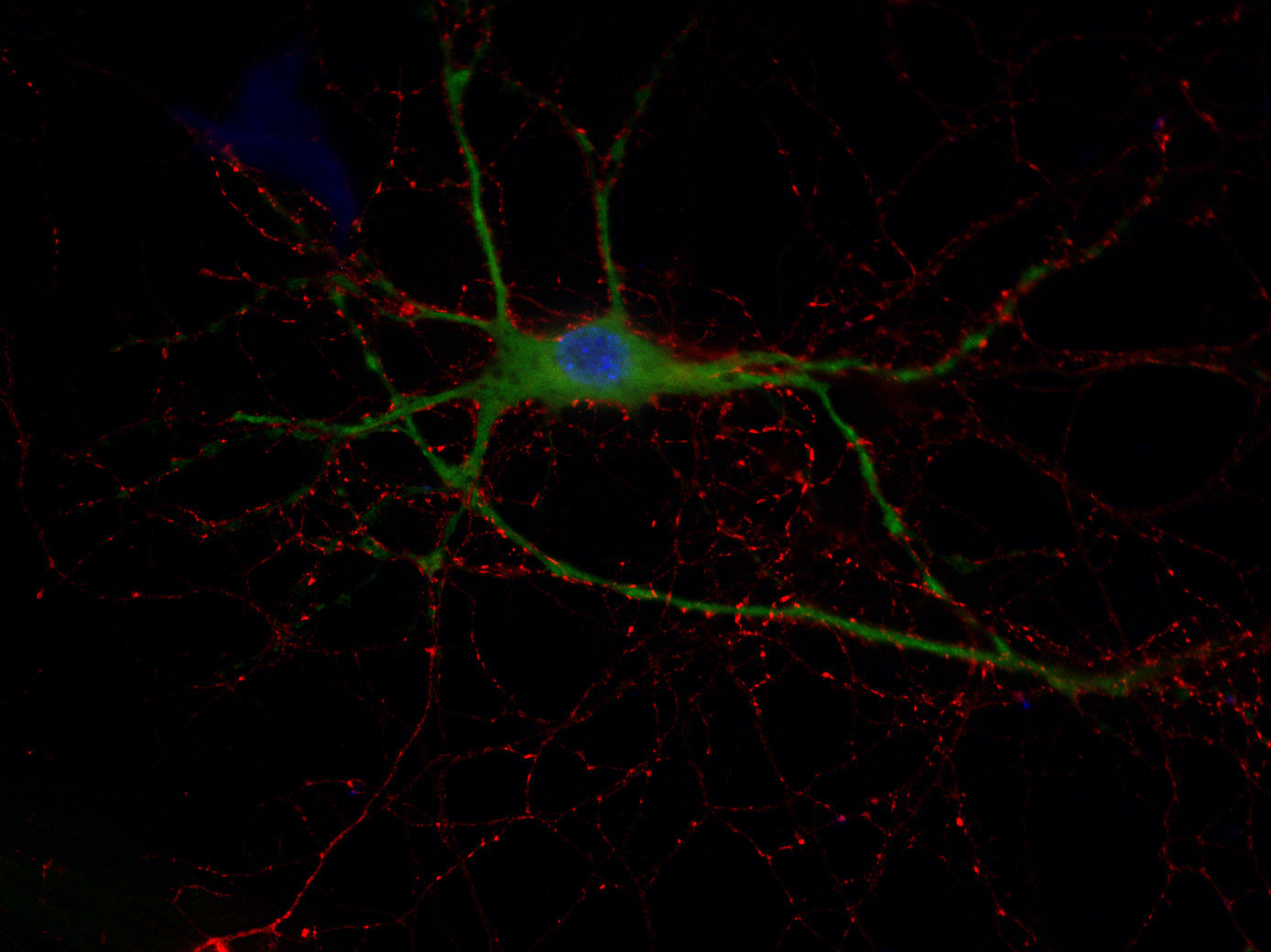
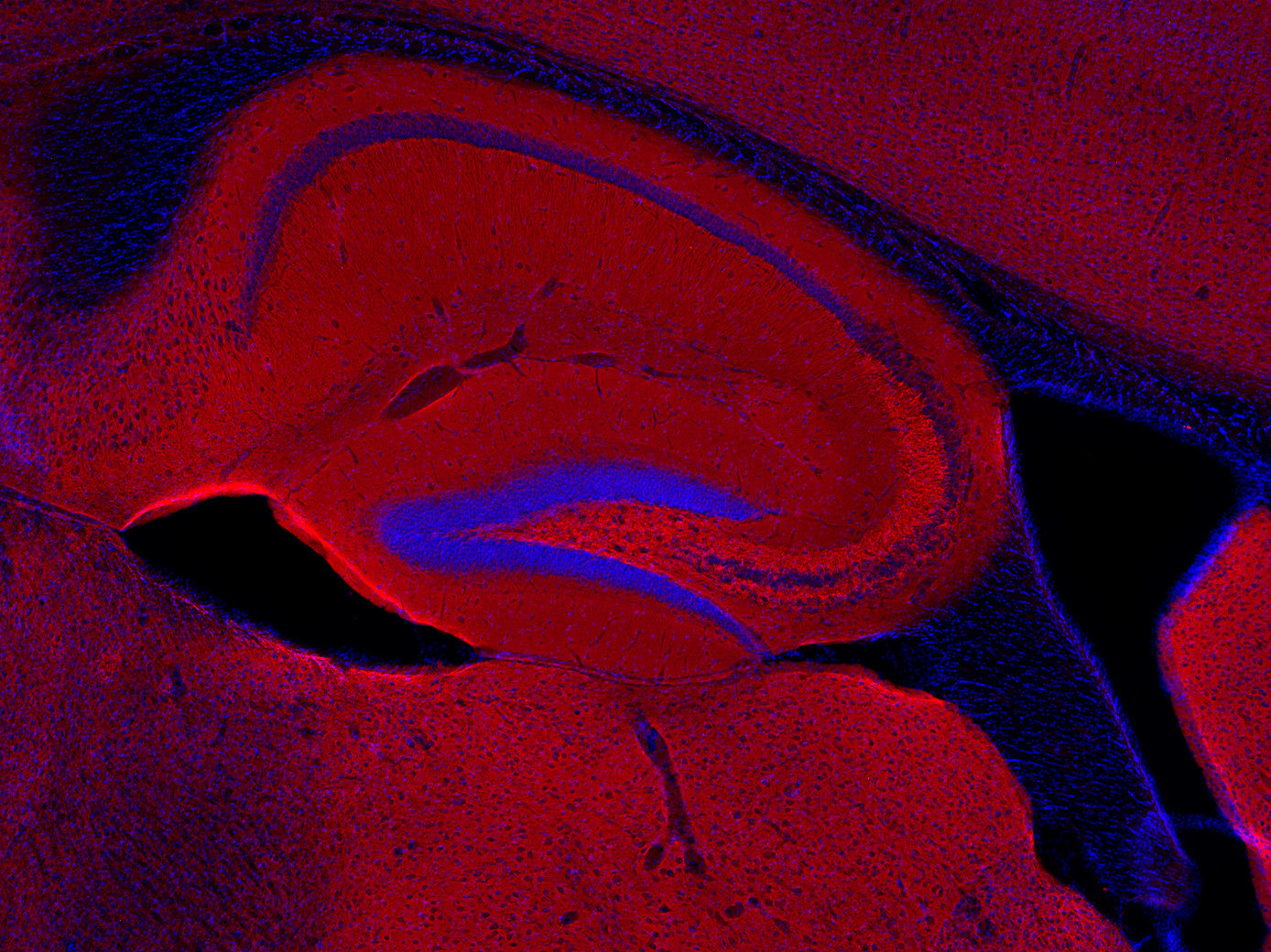
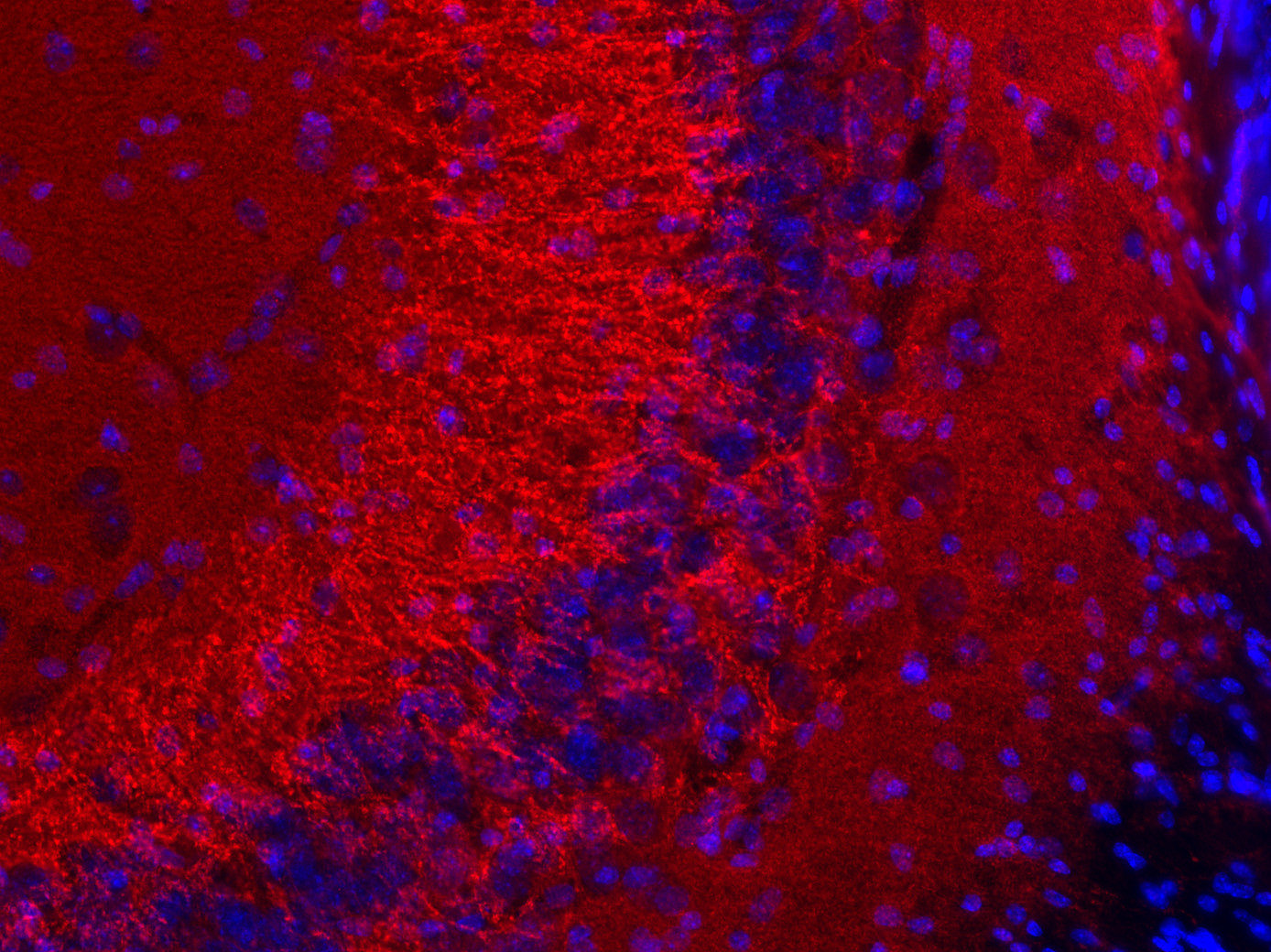
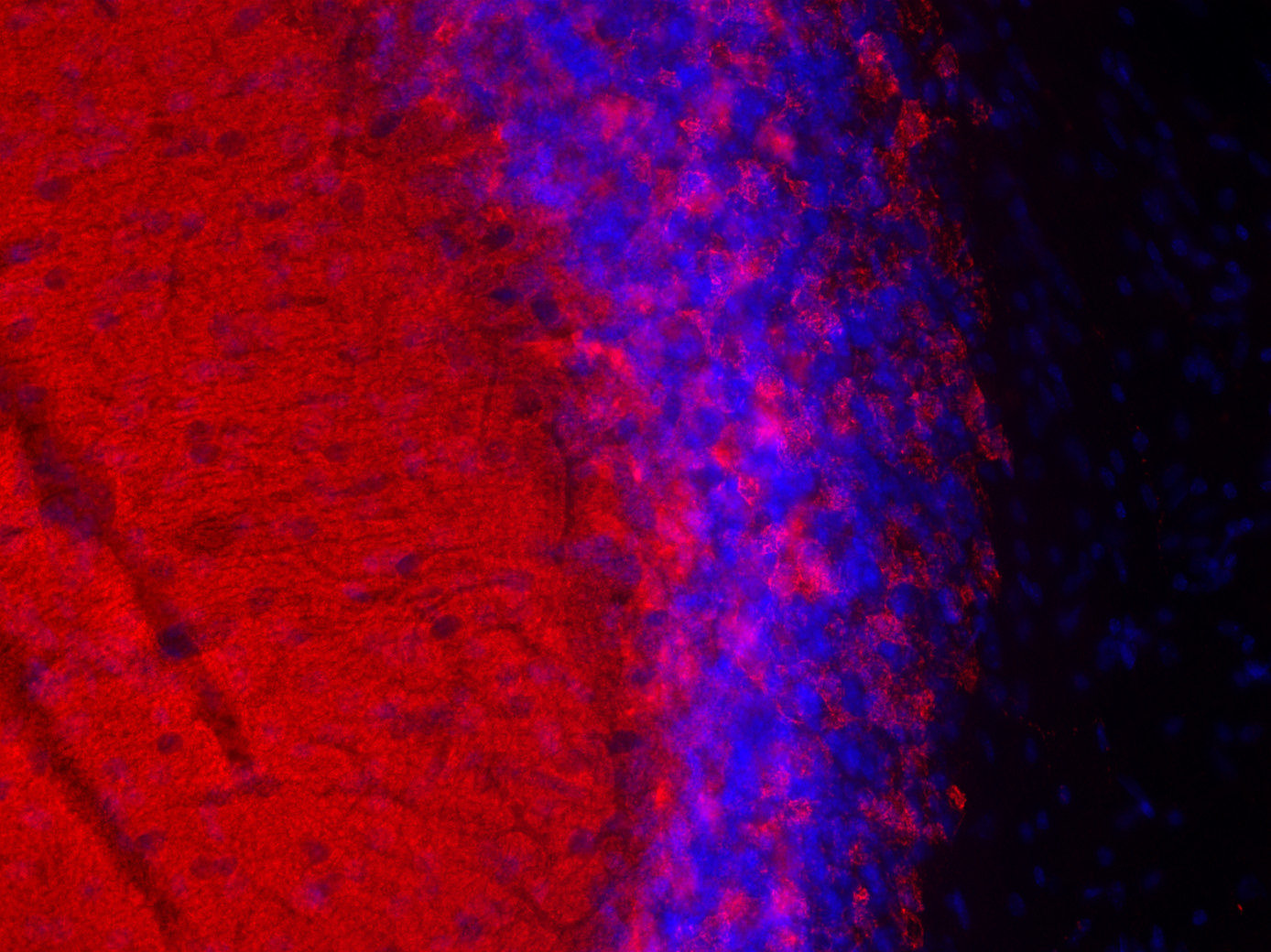
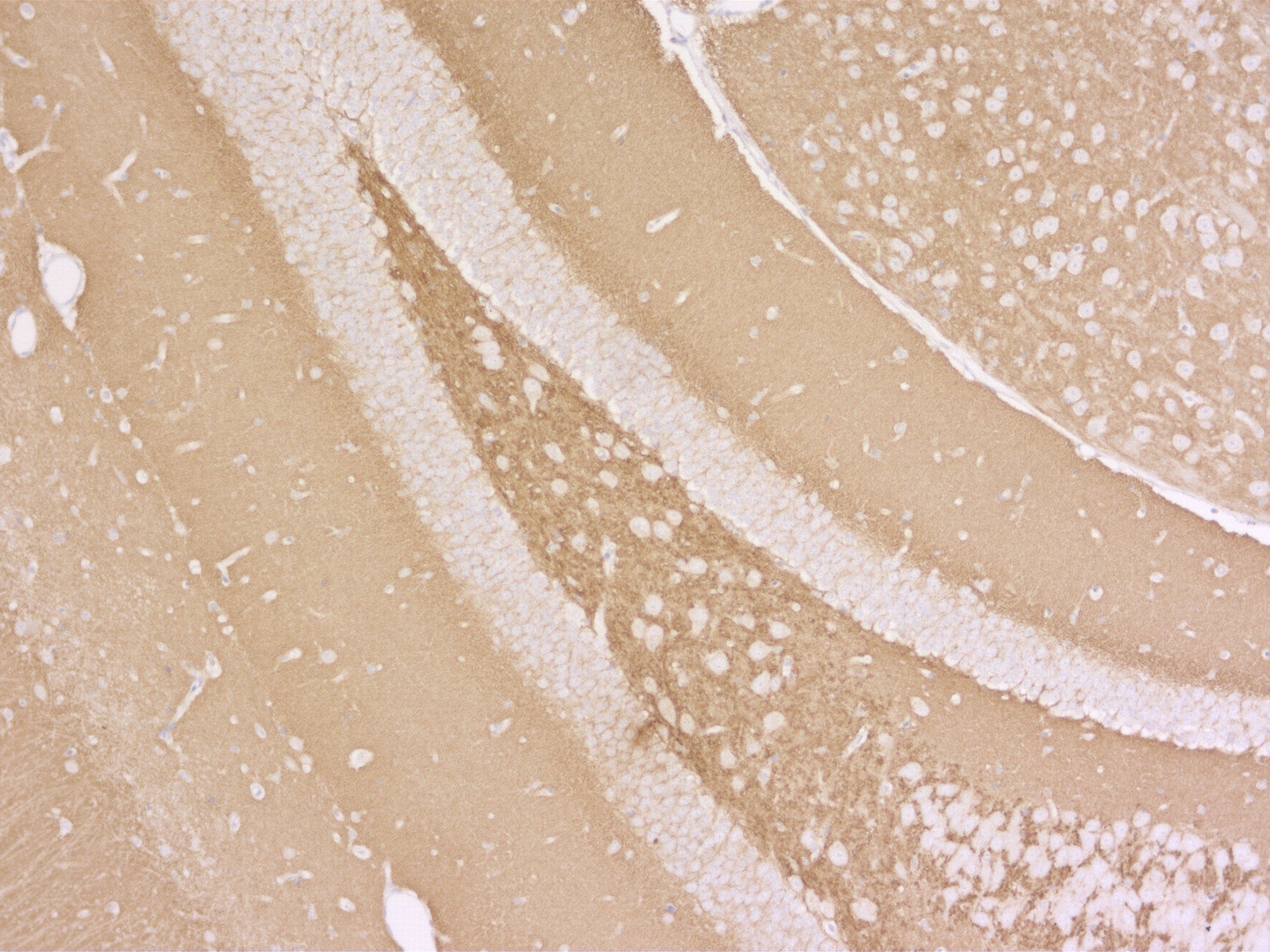
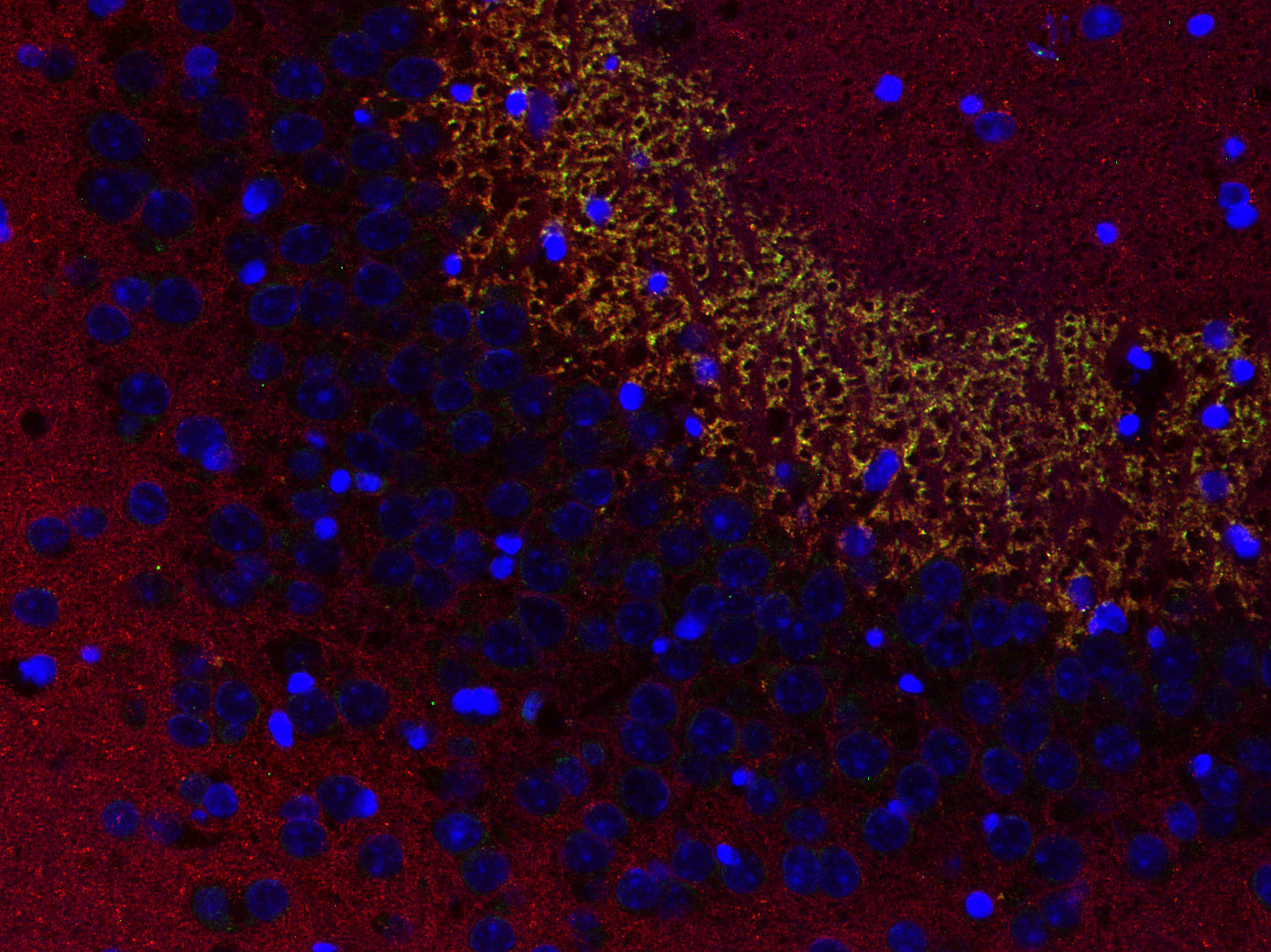
Synaptobrevins, also known as vesicle-associated membrane proteins (VAMPs), are predominantly expressed in the nervous system and are classified within the brevin subfamily of the SNARE (Soluble NSF Attachment Protein Receptor) protein superfamily. Brevins are small integral transmembrane proteins characterized by a central SNARE motif, an N-terminal cytoplasmic domain, and a C-terminal transmembrane domain. As crucial components of the SNARE machinery, these proteins play an essential role in vesicular transport and membrane fusion processes within cells (1, 2, 3).
In addition to synaptobrevins, the brevin family includes other tissue-specific members such as cellubrevin (VAMP3), myobrevin (VAMP5), and endobrevin (VAMP8), which are expressed in various non-neuronal tissues (4, 5, 6). These isoforms exhibit distinct spatial expression profiles, suggesting specialized functions beyond the nervous system.
Two Synaptobrevin isoforms were identified in the mammalian CNS, synaptobrevin1 (VAMP1 or p18-1) and synaptobrevin2 (VAMP2 or p18-2) that differ in their regional distribution within the brain, indicating isoform-specific roles in neuroexocytosis (7).
Synaptobrevin1 (VAMP1) is supposed to be essential for the maintenance of nerve impulse transmission in neuromuscular synapses. In addition, it is present on secretory granules of neuroendocrine cells. Synaptobrevin2 (VAMP2) is more abundant and widely distributed in the brain and has been shown to be mainly involved in the assembly of effective SNARE complexes, Ca2+-dependent SV exocytosis, and fast endocytosis in hippocampal synapses (8). It is also expressed in spinal cord dorsal horn neurons and implicated in inflammatory pain sensitization (9).
Synaptobrevins are target molecules for tetanus and several of the botulinal neurotoxins which cleave the protein at single sites in the C-terminal portion of the molecule and thereby disrupt neurotransmitter release (10).